Project A.06
- PhD student: Jessica Leismann
- Supervisor: Joan Barau
- Co-Supervisors: René Ketting
- Further TAC-members: Petra Beli
- Research Group
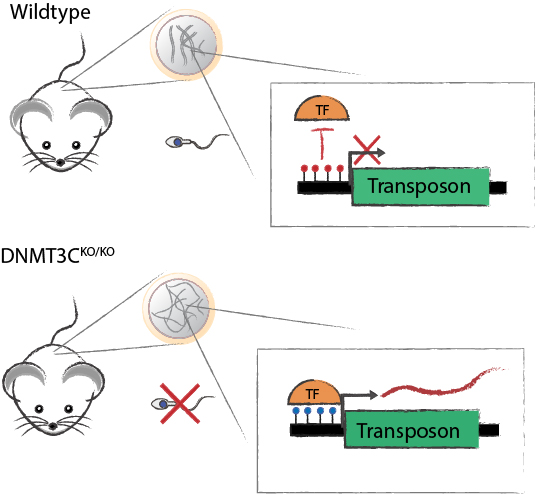
Transposons are abundant mobile DNA elements that pose a threat to genome stability and are therefore repressed by DNA methylation. In germline development, DNA methylation is erased allowing for activation of transposons, which is counteracted by the piRNA pathway methylating active transposons. Failure of this leads to reactivation of transposons later in development. This raises the question, what activates transposons in the absence of DNA methylation at meiosis and what represses transposons before.
Transposable elements (TEs) are highly abundant mobile units of DNA found in all kingdoms of life. Their activity can lead to biological innovation across evolutionary time scales. However, it also constitutes a source of genomic instability. Therefore, it is crucial that TEs are properly repressed. DNA methylation is a potent repressor of TEs when deposited at promoters.
During mammalian development DNA methylation undergoes two extensive waves of genome-wide reprogramming. A first, embryonic wave erases the gametic methylation patterns. A second, germline wave erases the somatic patterns derived from the developing embryo, and allows the de-repression of germline genes required for meiosis and gametogenesis. While DNA methylation reprogramming is very important for development, it also puts the genome at danger of instability due to de-repression of TEs. In fetal germ cells, this danger is neutralized by a genome defense pathway and de novo methylation in males, but not females. In this pathway, PIWI proteins detect and convert mRNAs from active TEs into small RNAs called piwi-interacting RNAs (piRNAs) and guide DNA methylation to repress active TE promoters. The methylation of TE promoters in the germline is performed by a muroid-specific DNA methyltransferase, DNMT3C, together with DNMT3L as a co-factor. In mice carrying a mutation for one gene of the genome defense pathway, failure to methylate TE promoters leads to the later reactivation of TEs at the onset of meiosis. As a result, these mutants are all male-sterile due to perturbations of meiotic functions1.
Interestingly, in pre-meiotic spermatogonia TE silencing occurs independently of DNA methylation. However, at the onset of meiosis, DNA methylation becomes essential for TE repression. What prevents the genotoxic TE reactivation in the hypomethylated genome of pre-meiotic cells and what leads to TE activation at meiosis remains unknown.
In my PhD project, I characterize and study the germline readers of DNA and chromatin methylation at TE promoters to uncover novel layers of regulation that protect the heritable genome from TEs, and reveal general principles of the epigenetic regulation of gametogenesis.